Ecological Biogeography
We've seen how energy and matter move through ecosystems. But if we want to fully understand ecosystems, we'll also need to look at ecological biogeography, which examines the distribution patterns of plants and animals from the viewpoint of their physiological needs. That is, we must examine how the individual organisms of an ecosystem interact with their environment. From fungi digesting organic matter on a forest floor to ospreys fishing in a coastal estuary, each organism has a range of environmental conditions that limits its survival as well as a set of characteristic adaptations that it exploits to obtain the energy it needs to live.
Let's start by looking at the relationship between organisms and their physical environment. Figure 8.12 shows how living conditions can change across the Canadian boreal forest such that different regions support different ecosystems. In this way, we can distinguish six distinct habitats across the Canadian boreal forest: upland, bog, bottomland, ridge, cliff, and active sand dune.
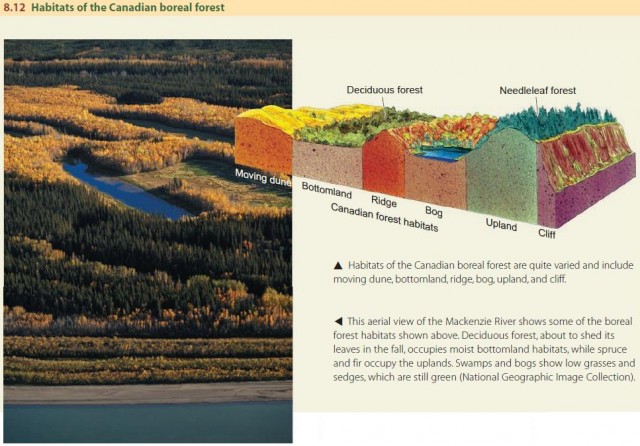
We use the term ecological niche to describe the functional role played by an organism as well as the physical space it inhabits. If the habitat is the individual's “address,” then the niche is its “profession,” including how and where it obtains its energy and how it influences other species and the environment around it.
When describing the ecological niche, we talk about the organism's tolerances and responses to changes in moisture, temperature, soil chemistry, illumination, and other factors. Although many different species may occupy the same habitat, only a few of these species will ever share the same ecological niche, for, as we'll see shortly, evolution will tend to separate those that do. As we move from habitat to habitat, we find that each is the home of a group of organisms that occupy different but interrelated ecological niches. We can define a community as an assemblage of organisms that live in a particular habitat and interact with one another.
Although every organism must adjust to variations in the environment on its own, we find that similar habitats often contain similar communities. Biogeographers and ecologists recognize specific types of communities, called associations, in which typical organisms are likely to be found together. These associations are usually defined by species, as in the beech-birch-maple forest that is found from the Great Lakes region to New England in suitable habitats.
WATER NEED
Let's now turn to the environmental factors that help determine where organisms, as individuals and species, are found. The first of these is the availability of water.
Plants and animals have adapted to cope with the abundance or scarcity of water in a variety of ways. Plants that are adapted to drought conditions are called xerophytes. (“Xerophyte” comes from the Greek roots xero-, meaning “dry,” and phyton, meaning “plant.”)
Some xerophytes have a thick layer of wax or waxlike material on leaves and stems, helping them to seal water inside. Others adapt to a desert environment by greatly reducing their leaf area or by bearing no leaves at all. Needlelike leaves, or spines in place of leaves, also conserve water.
Plants in water-scarce environments are also better at obtaining and storing water. For example, their roots may extend deeply to reach soil moisture far from the surface until they reach ground water. Plants drawing from ground water are called phreatophytes. Other desert plants produce a widespread, but shallow, root system, so they can absorb water from short desert downpours that saturate only the uppermost soil layer. Leaves and stems of desert plants known as succulents are often thickened by a spongy tissue that stores water. The common prickly pear cactus is an example.
Many small desert plants have a very short life cycle— germinating from seed, leafing out, bearing flowers, and producing seed in the few weeks immediately following a heavy rain shower. They survive the dry period as dormant seeds that require no moisture.
Certain climates, such as the wet-dry tropical climate 3 , have a yearly cycle with one season in which water is unavailable to plants because of lack of precipitation. In these climates, some species of trees, termed tropophytes, are deciduous, shedding their leaves at the onset of the dry season and growing new ones with the arrival of the wet season. The Mediterranean climate 7 also has a strong seasonal wet-dry alternation, with dry summers and wet winters. Plants in this climate often have hard, thick, leathery, evergreen leaves and are referred to as sclerophylls.
Xeric animals have evolved methods that are somewhat similar to those used by the plants. Many of the invertebrates stay dormant during the dry period. When rain falls, they emerge to take advantage of the new and short-lived vegetation that often results. Many species of birds only nest when the rains occur, the time of most abundant food for their offspring. The tiny brine shrimp of the Great Basin may wait many years in dormancy until normally dry lakebeds fill with water, an event that occurs perhaps three or four times a century. The shrimp then emerge and complete their life cycles before the lake evaporates. Other animals have evolved more unique adaptations, such as changing their body color to absorb or reflect solar energy, depending on their internal temperature.
Mammals are by nature poorly adapted to desert environments, but many survive through a variety of mechanisms that enable them to avoid water loss. Just as plants reduce transpiration to conserve water, many desert mammals do not sweat through skin glands. Instead they rely on other methods of cooling, such as avoiding the Sun and becoming active only at night. In this respect, they are joined by most of the rest of the desert fauna, which spend their days in cool burrows in the soil and their nights foraging for food.
TEMPERATURE
The temperature of the air and soil directly influences the rates of physiological processes in plant and animal tissues. In general, each plant species has an optimum temperature associated with each of its functions, such as photosynthesis, flowering, fruiting, or seed germination. There are limiting lower and upper temperatures for these individual functions as well and for the total survival of the plant itself.
Temperature can also act indirectly on plants and animals. Higher air temperatures reduce the relative humidity of the air, enhancing transpiration from plant leaves as well as increasing direct evaporation of soil water. In general, the colder the climate, the fewer the species capable of surviving. We only find a few plants and animals in the severely cold arctic and alpine environments of high latitudes and high altitudes. In plants, ice crystals can grow inside cells in freezing weather, disrupting cellular structures. Cold-tolerant plant species are able to expel excess water from cells to spaces between cells, where freezing does no damage.
Most animals can't regulate their temperature internally. These animals, including reptiles, invertebrates, fish, and amphibians, are cold-blooded animals—their body temperature passively follows the environment. With a few exceptions (notably fish and some social insects), these animals are active only during the warmer parts of the year. They survive the cold weather of the midlatitude zone winter by becoming dormant.
Some vertebrates enter a state called hibernation, in which their metabolic processes virtually stop and their body temperatures closely parallel those of the surroundings. Most hibernators seek out burrows, nests, or other environments where winter temperatures do not reach extremes or fluctuate rapidly. Soil burrows are particularly suited to hibernation because below the uppermost layers, soil temperatures don't vary a great deal.
Warm-blooded animals, like us, maintain tissues at a constant temperature by internal metabolism. This group includes the birds and mammals. Fur, hair, and feathers insulate the animals by trapping dead air spaces next to the skin surface. A thick layer of fat will also provide excellent insulation. Other adaptations are for cooling—for example, sweating or panting uses the high latent heat of vaporization of water to remove heat. The seal's flippers and bird's feet expose blood-circulating tissues to the cooler surroundings, promoting heat loss.
OTHER CLIMATIC FACTORS
Light also helps determine local plant distribution patterns. Some plants are adapted to bright sunlight, whereas others require shade. The amount of light available to a plant will depend in large part on the plant's position. Tree crowns in the upper layer of a forest receive maximum light but correspondingly reduce the amount available to lower layers. In extreme cases, forest trees so effectively cut off light that the forest floor is almost free of shrubs and smaller plants.
In certain deciduous forests of midlatitudes, the period of early spring, before the trees are in leaf, is one of high light intensity at ground level, permitting the smaller plants to go through a rapid growth cycle. In summer these plants largely disappear as the tree leaf canopy is completed. Other low plants in the same habitat require shade and do not appear until the leaf canopy is well developed.
The light available for plant growth varies by latitude and season. As we saw earlier, the number of daylight hours in summer increases rapidly with higher latitude and reaches its maximum poleward of the Arctic and Antarctic Circles, where the Sun may be above the horizon for 24 hours. The rate of plant growth in the short frost-free summer is greatly accelerated by the prolonged daylight.
In midlatitudes, where many species are deciduous, the annual rhythm of increasing and decreasing periods of daylight determines the timing of budding, flowering, fruiting, and leaf shedding. Even on overcast days there is usually enough light for most plants to carry out photosynthesis at their maximum rates.
Light also influences animal behavior. The day–night cycle controls the activity patterns of many animals. Birds, for example, are generally active during the day, whereas small foraging mammals, such as weasels, skunks, and chipmunks, are more active at night. In midlatitudes, as autumn days grow shorter and shorter, squirrels and other rodents hoard food for the coming winter season. Later, increasing hours of daylight in the spring trigger such activities as mating and reproduction.
Wind is also an important environmental factor in the structure of vegetation in highly exposed positions. Wind causes excessive drying, desiccating the exposed side of the plant and killing its leaves and shoots. Trees of high-mountain summits are often distorted in shape, with trunks and branches bent to near-horizontal, facing away from the prevailing wind direction.
Taken separately or together, moisture, temperature, light, and wind can limit the distribution of plant and animal species. Biogeographers recognize that there is a critical level of climatic stress beyond which a species cannot survive. This means that we can mark out a bioclimatic frontier—a geographic boundary showing the limits of the potential distribution of a species.
GEOMORPHIC FACTORS
Geomorphic, or landform, factors such as slope steepness, slope aspect (the orientation of a sloping ground surface with respect to geographic north), and relief (the difference in elevation of divides and adjacent valley bottoms) help differentiate habitats for ecosystems.
Slope steepness affects the rate at which precipitation drains from a surface, which indirectly influences plants and animals. On steep slopes, surface runoff is rapid, but on gentle slopes, more precipitation penetrates into the soil, providing a moister habitat. Steep slopes often have thin soil because they are more easily eroded, while soil on gentler slopes is thicker. Slope aspect controls plants' exposure to sunlight and prevailing winds. Slopes facing the Sun have a warmer, drier environment than slopes that face away from the Sun. In midlatitudes, these slope-aspect contrasts may be strong enough to produce quite different biotic communities on north-facing and south-facing slopes.
On divides, peaks, and ridge crests, rapid drainage dries the soil, which is also more exposed to sunlight and drying winds. By contrast, the valley floors are wetter because water converges there. In humid climates, the ground water table in valley floors may lie close to or at the ground surface, producing marshes, swamps, ponds, and bogs.
EDAPHIC FACTORS
Soils can vary widely from one small area to the next, influencing the local distribution of plants and animals. Edaphic factors are connected to the soil. For example, sandy soils store less water than soils with abundant silt and clay, so they are often home to xerophytes. If there's a high amount of organic matter in the soil, then the soil will be rich in nutrients and will harbor more plant species. The relationship can work in the opposite direction, too—biota can change soil conditions, as when a prairie grassland builds a rich, fertile soil beneath it.
DISTURBANCE
Disturbance includes fire, flood, volcanic eruption, storm waves, high winds, and other infrequent catastrophic events that damage or destroy ecosystems and modify habitats. Although disturbance can greatly alter the nature of an ecosystem, it is often part of a natural cycle of regeneration that gives short-lived or specialized species the opportunity to grow and reproduce.
For example, fire will strike most forests sooner or later. In many cases, the fire is beneficial. It cleans out the understory and consumes dead and decaying organic matter while leaving most of the overstory trees untouched.
Fire helps expose mineral soil on the forest floor and fertilizes it with new ash, providing a productive environment for dormant seeds. In addition, shrubs and forbs no longer shade the soil from sunlight. Among tree species, pines are typically well adapted to germinating under such conditions. In fact, the jack pine of eastern North America and the lodgepole pine of the intermountain West have cones that remain tightly closed until the heat of a fire opens them, allowing the seeds to be released.
Fires also preserve grasslands. Grasses are fire-resistant because they have extensive root systems below ground and germinal buds located at or just below the surface. But woody plants that might otherwise invade grassland areas are not so resistant and are usually killed by grass fires. In many regions, active fire suppression has reduced the frequency of burning to well below natural levels. That may sound like a good thing, but in forests, this causes dead wood to build up on the forest floor. So, when a fire does start, it's destructive—burning hotter and more rapidly and consuming the crowns of many overstory trees.
Flooding is another important disturbance. It displaces animal communities and also deprives plant roots of oxygen. Where flooding brings a swift current, mechanical damage rips limbs from trees and scours out roots. High winds are another significant factor that can topple individual trees as well as whole forest stands.
INTERACTIONS AMONG SPECIES
Species don't react with just their physical surroundings. They also interact with each other. That interaction may benefit at least one of the species, be negative to one or both species, or have no effect on either species.
Competition is a negative interaction. It happens whenever two species need a common resource that is in short supply. Both populations suffer from lowered growth rates than they would have had if only one species were present. Sometimes one species will win the competition and crowd out its competitor. At other times, the two species may remain in competition indefinitely.
Competition is an unstable situation. If a genetic strain within one of the populations emerges that can use a substitute resource, its survival rate will be higher than that of the remaining strain, which still competes. The original strain may become extinct. In this way, evolutionary mechanisms tend to reduce competition among species.
Predation and parasitism are other negative interactions between species. Predation occurs when one species feeds on another. There are obvious benefits for the predator species, which obtains energy for survival, but, of course, the interaction has a negative outcome for the prey species. Parasitism occurs when one species gains nutrition from another, typically when the parasite organism invades or attaches to the body of the host in some way.
Although we tend to think that predation and parasitism are always negative—benefiting one species at the expense of the other—in some cases it works out well for the prey or host populations, too, in the long run. A classic example is the rise and fall of the deer herd on the Kaibab Plateau north of the Grand Canyon in Arizona. Predation and parasitism will also remove the weaker individuals, improving the attacked species' genetic composition.
Another type of negative interaction between species is herbivory. When animals graze, they can reduce the viability of the plant species population. Although some plants can maintain themselves well in the face of grazing pressure, others are quite sensitive to overgrazing. Allelopathy, also a negative interaction, occurs when one plant species produces chemical toxins that inhibit other species.
We mentioned the symbiotic relationship between legumes and the nitrogen-fixing Rhizobium bacteria— which benefits both species—when we looked at the nitrogen cycle, earlier in the chapter. Symbiosis includes three types of positive interactions: commensalism, protocooperation, and mutualism. In commensalism, one of the species is benefited and the other is unaffected. Sometimes the relationship benefits both parties but isn't essential for their survival. This type of relationship is called protocooperation. If the relationship reaches a point where one or both species cannot survive alone, it's called mutualism. The relationship between the nitrogen-fixing bacterium Rhizobium and legumes is a classic example of mutualism because Rhizobium needs the plant for its own survival.